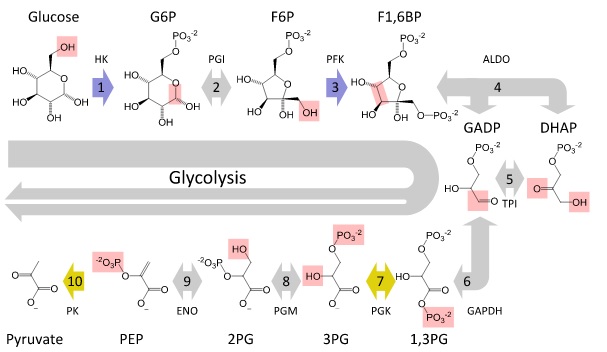
Этапы расписаны ниже:
- Вы начинаете с глюкозы, проходящей через канал белка из-за градиента концентрации.
- Затем глюкоза необратимо фосфорилируется с использованием гексокиназы (HK) и одного АТФ с образованием глюкозо-6-фосфата (G6P), поскольку кислород шестого углерода является осевым и, следовательно, к нему проще всего получить доступ. Это необходимо, потому что на следующем этапе есть фермент, который требует фосфорилированной глюкозы.
- Затем G6P необходимо изомеризовать с помощью фосфоглюкозоизомеразы (PGI) с образованием фруктозо-6-фосфата (F6P), поскольку цель состоит в том, чтобы получить два 3-углеродных соединения, которые могут находиться в равновесии друг с другом (дигидроксиацетон и L-глицеральдегид). Это шаг равновесия.
- Затем F6P необходимо фосфорилировать в положении 1 с использованием субъединицы PFK-1 в комплексе фосфофруктокиназы и одного АТФ, чтобы после получения дигидроксиацетона и L-глицеральдегида они оба были фосфорилированы. Вы получаете фруктозу-1,6-ВР (F16BP).
Это первый совершенный шаг и необратимый.
- F16BP затем может быть расщеплен с использованием альдолазы с образованием двух 3-углеродных соединений, о которых я говорил ранее: дигидроксиацетонфосфат (DHAP) и глицеральдегид-3-фосфат (GAP).
- DHAP и GAP находятся в равновесии друг с другом. Триозофосфатизомераза (TIM) изомеризует DHAP в GAP . Это равновесие продвигается вперед от производства DHAP из-за предыдущих шагов.
- ВАЖНЫЙ ШАГ: Два эквивалента GAP используются с2 NAD +, две свободные фосфорильные группы (2 P i), и фермент глицеральдегид-3-фосфатдегидрогеназы (GAPDH) вместе со специальным механизмом, который дает вам высокоэнергетический тиоэфирный промежуточный продукт, чтобы дать вам два 1,3-бисфосфоглицерата ( 2 1,3-BPG) и2 НАД,
На этом этапе вводятся две свободные фосфорильные группы, что в конечном итоге означает, что вы получили два АТФ-эквивалента. Это то, что позволяет вам получить чистую прибыль в ATP.
- Затем 1,3-BPG использует фосфоглицераткиназу (которая противоречит интуитивному принципу , но ее называют обратной реакцией) для дефосфорилирования одного положения и получения 3-фосфоглицерата (3-PG).
При этом используется один эквивалент ADP для создания одного эквивалента ATP.
- Как только мы здесь, 3-PG преобразуется в 2-PG с использованием фермента, называемого фосфоглицерат мутаза (PGM). Это проходит через промежуточный 2,3-BPG (который вы, возможно, видели, связывается с центральной полостью в гемоглобине).
- 2-PG теперь должен стать енолом, использующим енолазу для образования фосфоенолпирувата (PEP). Он нуждается в ферменте, поскольку обычно энергетически не выгодно образовывать енолят из-за отсутствия эффективного резонанса.
- Наконец, PEP превращается в пируват с использованием пируваткиназы , высвобождая энергию. Этот шаг необратим из-за этой высвобождаемой энергии.
При этом используется один эквивалент ADP для создания одного эквивалента ATP.
В целом, вы использовали 1 эквивалент глюкозы, чтобы получить 2 эквивалента пирувата, использовали 2 эквивалента АТФ и получили 4 эквивалента АТФ, использовали 2 эквивалента NAD + и получили 2 эквивалента NADH.









Новые комментарии